
在人类细胞中同时精确编辑多个基因及筛选单克隆细胞的方法
作者:艾贝泰 2022.07.29 点击601次
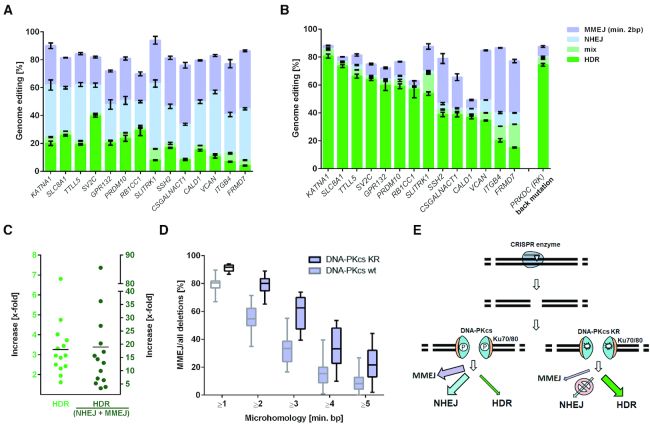
CRISPR技术的发现带来了一种高效、可靠、方便的基因组编辑工具的希望。但是,过去用Cas9靶标多个基因组位点需要构建多个或者很大的表达载体,CRISPR-Cas9编辑技术的应用具有一定的局限性。而多重基因组编辑技术可以高精度的修改基因组中两个或多个特定的DNA位点,大大增加了在多个核苷酸水平上向目标基因组引入突变的可行性。 CRISPR核酸酶和向导RNA(gRNA)被用来在基因组中引入双链断裂(DSB),基因组编辑的准确性和效率受到细胞DSB修复途径的影响,这些修复途径包括同源定向修复(HDR)、非同源端连接(NHEJ)、微同源介导的末端连接(MMEJ)等[1]。NHEJ是大多数真核生物修复DSB的主要细胞修复途径,但常常因能引入indel从而导致移码突变;而HDR通常使用姐妹染色单体作为同源修复模板来精准修复携带DNA断裂的染色体,从而实现精确的基因组编辑(PGE),但效率比较低。 德国马克斯普朗克进化人类学研究所研究人员Stephan Riesenberg等人分享一种基于CRISPR-Cas9的多重基因组编辑,利用PRKDC基因突变和激酶抑制剂M3814抑制DNA依赖性蛋白激酶的催化亚单位(DNA-PKcs)活性,增加基因编辑HDR效率,使得在同一个细胞中同时精确编辑多达4个基因的方法[2]。 人类细胞PRKDC基因的K3753R突变可促进HDR 为了提高靶向DSB的基因编辑效率,减少DSB的脱靶可能,建立人诱导多能干细胞(hiPSC)细胞系{在含Matrigel Matrix (Corning, 35248)的板中培养,培养基为每天更换补充剂 (StemCell Technologies, 05852) 的mTeSR1 培养基 (StemCell Technologies, 05851)},携带D10A突变的强力霉素诱导型Cas9(iCRISPR-Cas9n),使其诱导DNA单链断裂[3]。hiPSC引入编码DNA-PKcs蛋白的PRKDC基因的K3753R突变,致其失活,对比DNA-PKcs K3753R(KR)和DNA-PKcs野生型(WT)细胞中引入核苷酸替换的效率。 设计gRNAs和14种单链寡脱氧核苷酸供体(ssODNs),强力霉素诱导Cas9n表达,转染gRNAs和ssODN。培养3天后分离DNA,PCR扩增目标基因并测序,鉴定ssODNs通过HDR插入靶标基因的效率。当单独编辑每个基因时,WT细胞中的HDR频率在14个基因中变化在4%到40%之间(平均18%)(图1A);在KR细胞中,频率在15%到81%之间(平均51%)(图1B)。在这些基因中,KR细胞的HDR增加了1.6倍到6.8倍(平均3.3倍)(图1C)。在KR细胞中,MMEJ相对于NHEJ增加(图1D),NHEJ被抑制,导致HDR急剧增加(图1E)。
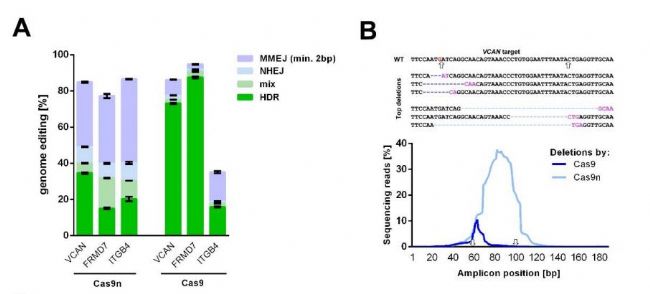
图1:失活DNA-PKcs可促进HDR 选取KR细胞中HDR频率最低的3个基因,使用Cas9替代Cas9n,其中两个基因的HDR分别从35%和20%增加到73%和87%(图2A)。因此,对于一些MMEJ占主导的靶点,可以通过使用不同的核酸酶提高PGE,以基因VCAN中Cas9n和Cas9编辑后的缺失模式为例,不同的切割位点,导致HDR频率的不同(图2B)。
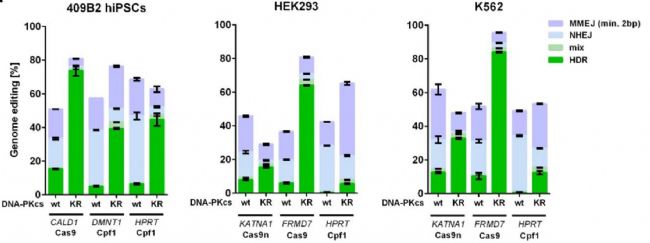
图2:不同核酸酶在KR细胞系中HDR频率变化 为了测试在hiPSCs中看到HDR的增加是否依赖于细胞类型或使用的酶,基因编辑HEK293细胞{(ECACC, 85120602),培养基为含 10% 胎牛血清 (FBS) (SIGMA, F2442) 和 1%NEAA(SIGMA, M7145)的Dulbecco 改良 Eagle 培养基/F-12 (Gibco, 31330-038)}和K562细胞{(ECACC, 89121407) ,培养基为含10% FBS的 Iscove 改良的 Dulbecco 培养基 (ThermoFisher, 12440053)},在单细胞打印机分选基因编辑后的单克隆细胞,进行文库制备和Illumina测序。分别对比在KR和WT中使用不同核酸酶Cas9n、Cas9和Cpf1的基因组编辑频率情况。可以发现在不同细胞系、不同核酸酶下,DNA-PKcs蛋白的PRKDC基因的K3753R突变都会引起HDR频率的增加(图3)。
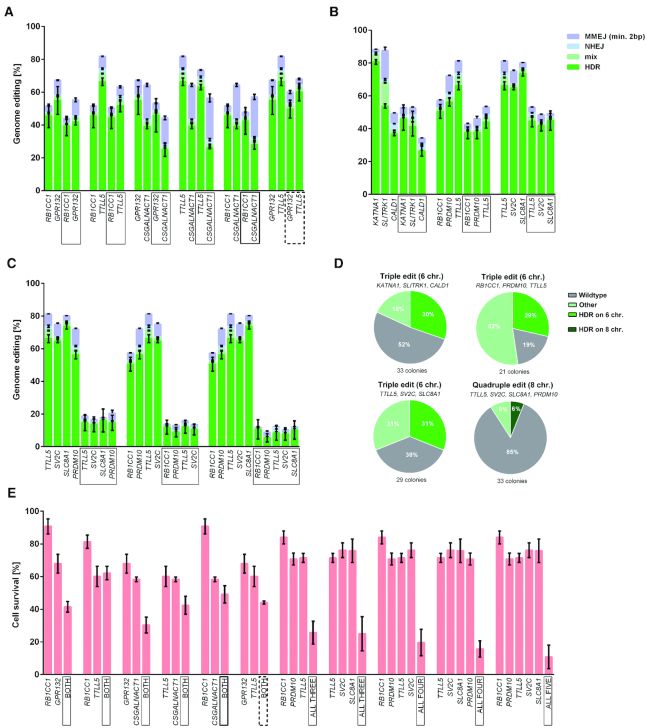
图3:不同DNA-PKcs K3753R细胞系能促进HDR 多个基因同时进行精确基因组编辑 将四种不同基因的成对组合在诱导Cas9n表达后电穿孔到iPSC的KR细胞中。409-B2 iCRISPR-Cas9n hiPSC 在编辑前三到四天在含有 2 g/ml 强力霉素 (Clontech, 631311) 的培养基中孵育。对于多重精确基因组编辑,在电穿孔前一天将该培养基更换为含有补充剂 (Gibco, A3349401)、CloneR (StemCell Technologies, 05888) 和强力霉素的 StemFlex。90% 的电穿孔细胞被铺板用于批量基因型分析,10% 的 hiPSC 被铺板在6 孔板中以产生源自单细胞的克隆,在电穿孔后 2 天,培养基中添加了 ROCK 抑制剂 Y-27632。至少 7 天后,挑选单克隆用于后续繁殖和 DNA 分离。当同时编辑两个基因时的HDR频率都与对应基因的单编辑相似或略低(图4A);对于三个基因的三种不同组合,HDR效率平均比单个编辑低三分之一,导致每个目标位点的平均编辑频率为40%(图4B);对于4个或5个基因,平均HDR频率分别下降到13%和8%(图4C)。此外,细胞存活率从2次编辑的30-68%下降到5次编辑的11%(图4E)。 对于三重编辑,分选的单细胞来源的克隆(SCCs)中大约有三分之一以纯合子的形式携带所有目标基因(6条染色体);四重基因编辑的SCCs中有6%的4个氨基酸替换的纯合子细胞克隆,比预期高出1000倍(图4D)。这表明,具有“编辑能力”的细胞往往会在多个位点上被有效编辑。
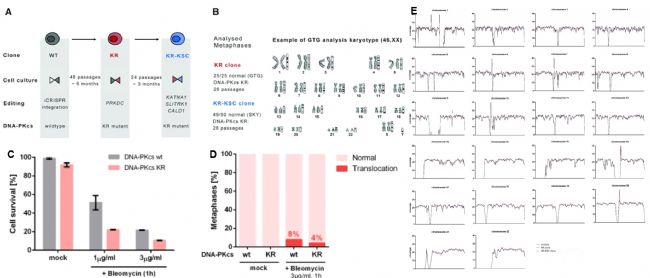
图4:利用失活DNA-PKcs细胞系进行多重精确基因组编辑 表达失活的DNA-PKcs的细胞的基因组稳定性不受影响 为了验证失活409B2-iCRISPR iPSC的DNA-PKcs催化活性是否导致基因组不稳定,使用Trypsin-Giemsa Banding(GTG)试剂和光谱核型分析(SKY)核型分析,显示KR-KCS克隆中观察到一个多倍体中期染色体(图5B)。此外,用DSB诱导药物博莱霉素处理表达WT和KR细胞,KR细胞比WT细胞的存活率降低高两倍(图5C)。经博莱霉素处理后,50个WT细胞中有4个中期染色体出现不平衡易位,50个KR细胞中有2个中期染色体出现平衡易位。因此,与WT细胞相比,KR细胞的非整倍体或染色体重排没有增加(图5D)。 为了评估其他类型的遗传不稳定性,对DNA-PKcs WT、WT细胞传代48次后生成KR克隆、KR细胞传代24次后生成三编辑克隆KR-KSC(KATNAI-SLITRKI-CALDI编辑)(图5A)进行全基因组测序。比较沿着染色体1mb基因组覆盖率,与WT相比,KR和KR-KSC的唯一差异是9号染色体覆盖率下降,表明存在杂合缺失(图5E)。全基因组测序结果发现在KR和KR- KSC细胞之间共有19个新的杂合单核苷酸变异(SNV),在KR- KSC细胞中有3个新的杂合SNV;没有发现三个细胞系之间的indel有什么不同。按全基因组结果推算,WT细胞传代的突变率为21,KR细胞为7,证明KR以及KR-KSC均没有提高突变的概率。
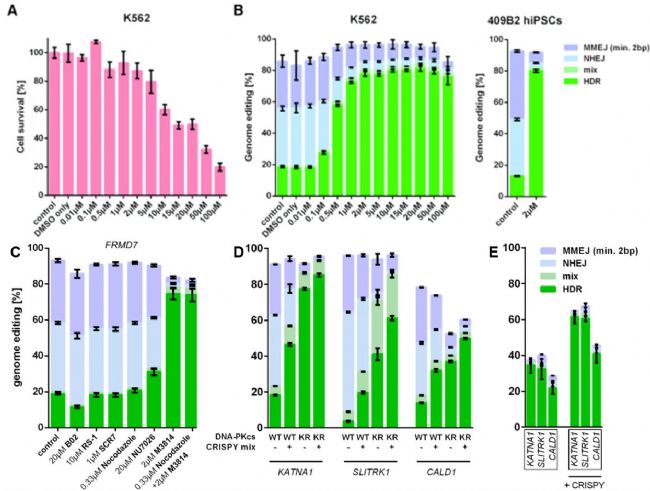
图5:DNA-PKcs K3753R细胞的基因组稳定性 小分子M3814瞬间失活DNA-PKcs 在KR细胞中提高核苷酸替换的效率,引入所需的改变后,恢复正常的DNA-PKcs功能成为可能,可以通过暂时抑制DNA-PKcs的激酶活性来增加HDR。M3814是一种新型高效的DNA-PKcs抑制剂,基因编辑的细胞在电转后分别使用或不使用2 μM的M3814处理3天后,用单细胞打印机(Cytena)分选单克隆细胞,提取DNA进行PCR扩增,制作Illumina文库并进行测序。 结果显示,在WT K562细胞中Cas9诱导的DSB的HDR从18%增加到81%,同时表现出中等毒性,409B2 hiPSCs的HDR也出现了类似的增加(图6A和6B)。与其他DNA-PK抑制剂相比,M3814使得HDR增加的效果更好(图6C)。为了探索这种明显的高编辑效率是否可能是由于诱导目标DNA序列的大量缺失,在无M3814和有M3814编辑后分离单克隆细胞,通过扩增子测序发现,当M3814未被使用时,10%的克隆以纯合子状态替换核苷酸;而使用M3814时,替换正确的克隆达到76%。另外,M3814同样增加了多重基因组编辑的HDR(图6D和6E)。因此,通过可逆性K3753R突变或M3814抑制DNA-PKcs的激酶活性,可以显著增加PGE,允许在人类细胞中同时引入多个核苷酸替换。
图6:小分子M3814对DNA-PKcs瞬时失活的影响 结 论 本篇文章展示人类细胞可以利用PRKDC基因的K3753R突变提高基因编辑中PGE的效率,并且可以和小分子M3814结合,进一步促进HDR。在这种方法下,没有发现K3753R突变会降低基因组稳定性;在博莱霉素处理后易位更少,每次传代的突变也更少。易出错的NHEJ的损伤可能会导致细胞通过不易出错的HDR更频繁地修复DSB,否则细胞可能会发生凋亡,导致存活的细胞以更准确的形式维持其基因组。DNA-PKcs抑制剂M3814能够在一定程度上增加HDR,与PRKDC突变可以达到相当的程度。永久性的DNA-PKcs失活会导致哺乳动物因无法进行V(D)J重组而产生严重的联合免疫缺陷,因此M3814可以在基因治疗中使用,提高HDR效率来实现治疗目标。
[1]. Xue C, Greene EC. DNA Repair Pathway Choices in CRISPR-Cas9-Mediated Genome Editing. Trends Genet. 2021 Jul;37(7):639-656. doi: 10.1016/j.tig.2021.02.008. Epub 2021 Apr 22. [2]. Riesenberg S, Chintalapati M, Macak D, Kanis P, Maricic T, Pääbo S. Simultaneous precise editing of multiple genes in human cells. Nucleic Acids Res. 2019 Nov 4;47(19):e116. doi: 10.1093/nar/gkz669. [3]. Riesenberg S, Maricic T. Targeting repair pathways with small molecules increases precise genome editing in pluripotent stem cells. Nat Commun. 2018 Jun 4;9(1):2164. doi: 10.1038/s41467-018-04609-7. | 产品分类
重点推荐产品
|






{kind=link}